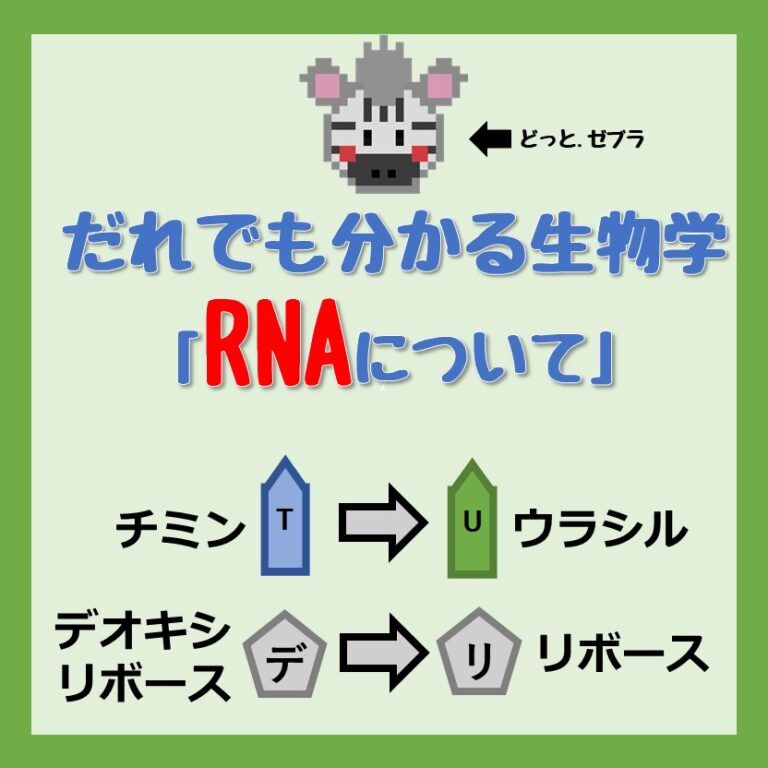
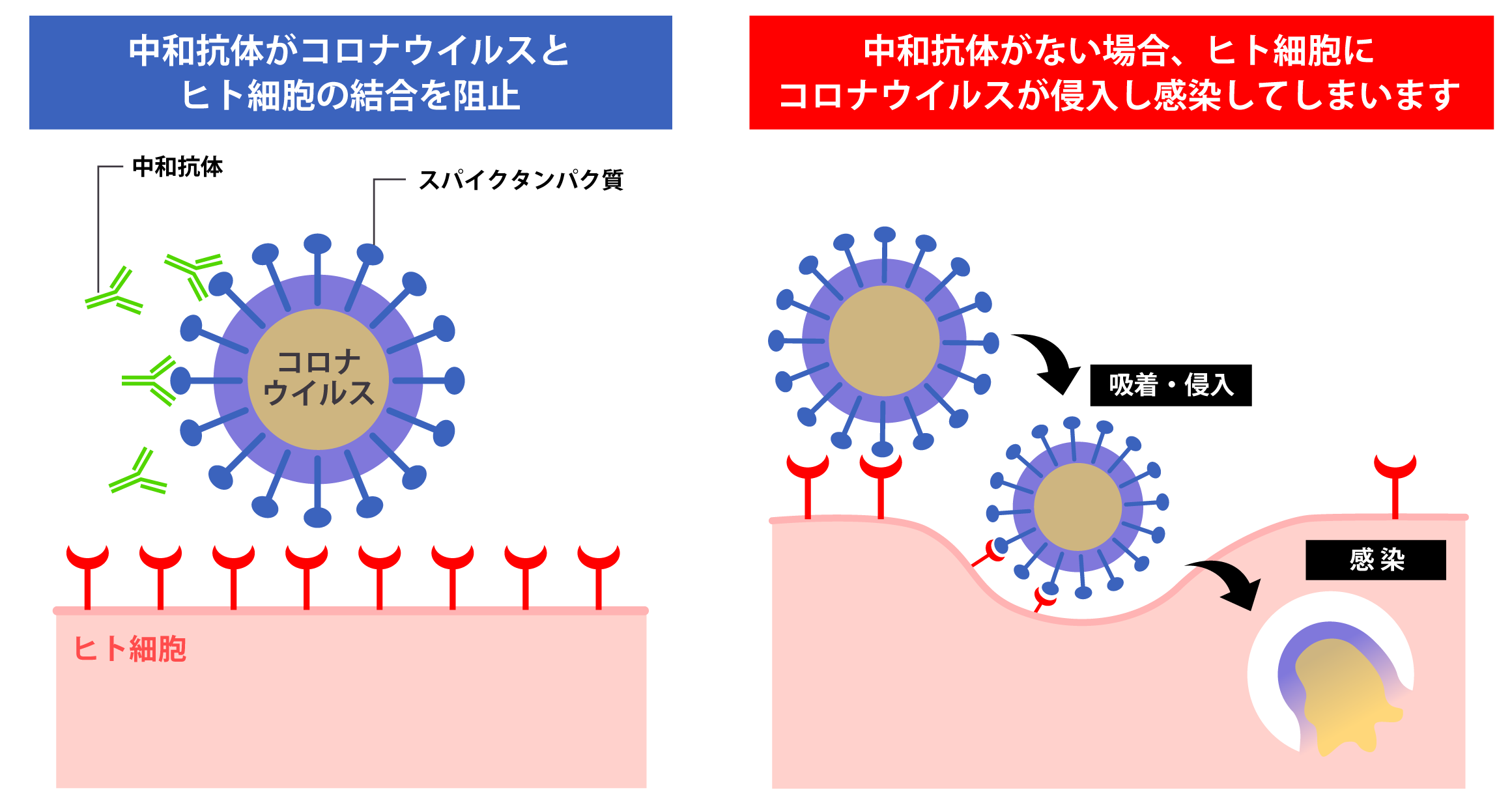
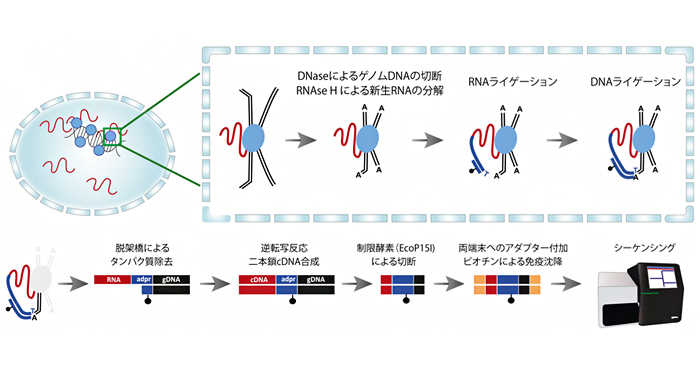
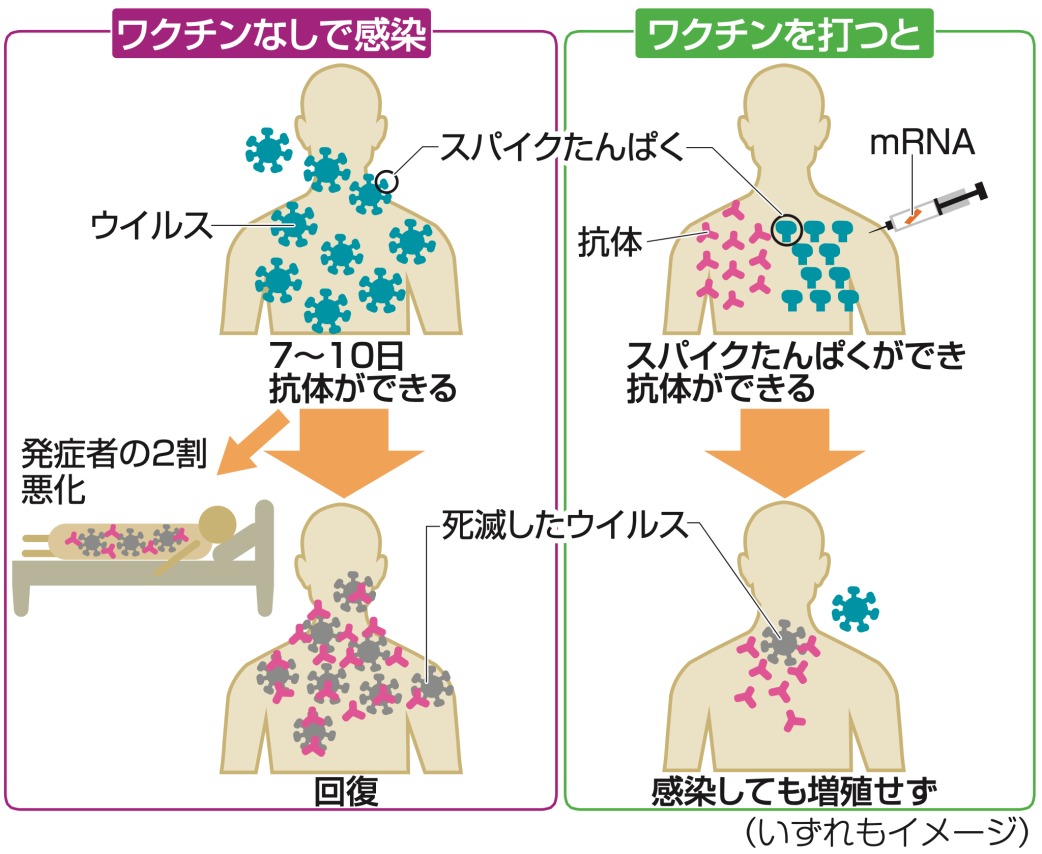
Images of RNA




![[ 分別ダストワゴン タワー / 蓋付き / 目隠し / 2分別 / 3分別 / 45L ]山崎実業 tower キッチン ゴミ箱 ごみ箱 ダストボックス ゴミ袋ハンガー フタ付き ふた 分別 ゴミ袋ホルダー キャスター付き レジ袋ホルダー 大容量 黒 白 おしゃれ カウンター下 北欧 公式](https://thumbnail.image.rakuten.co.jp/@0_mall/roomy/cabinet/500cart_all/500cart_11g/p5-7/ymz1002-st019-0_gt01.jpg?_ex=300x300)
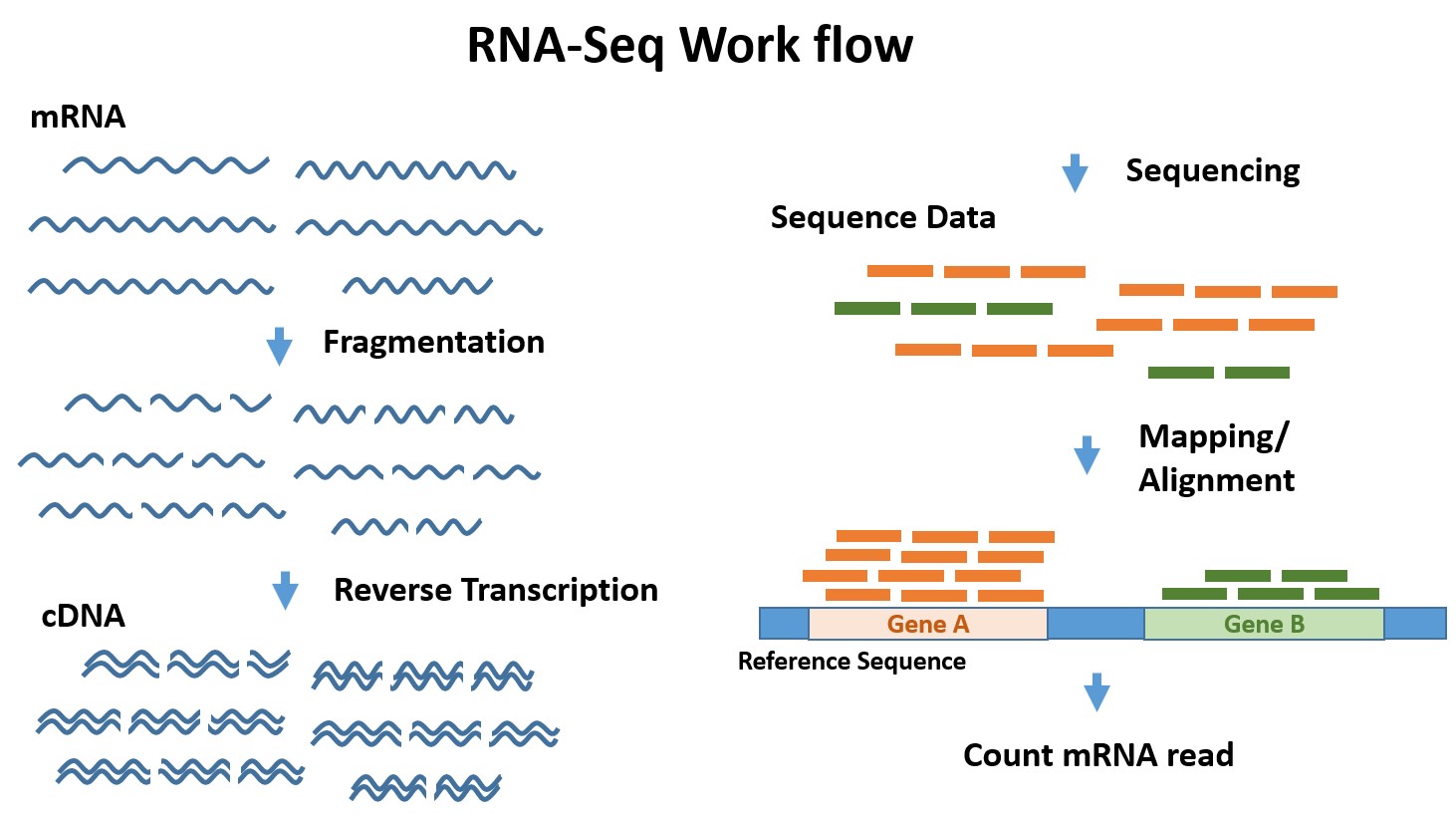
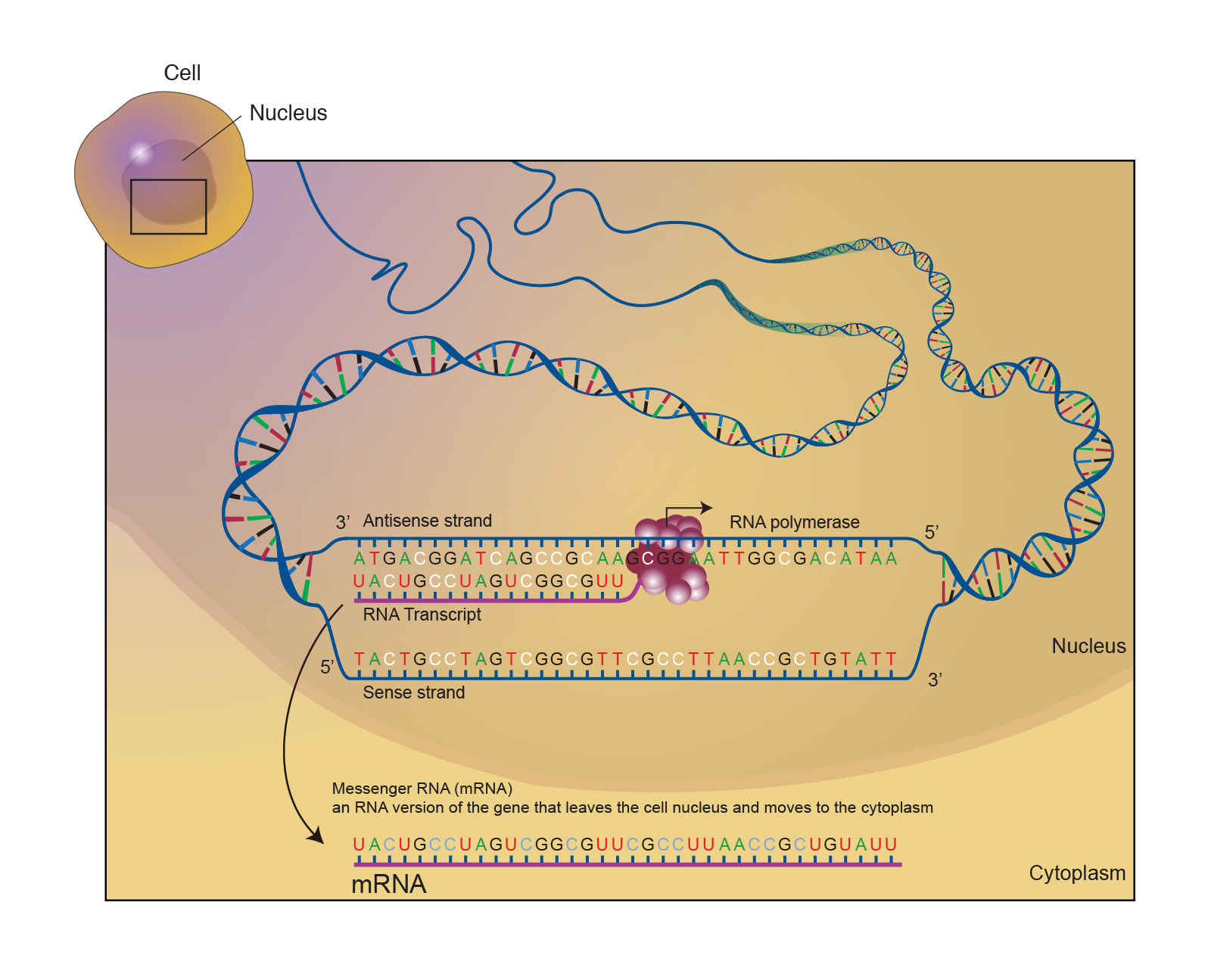

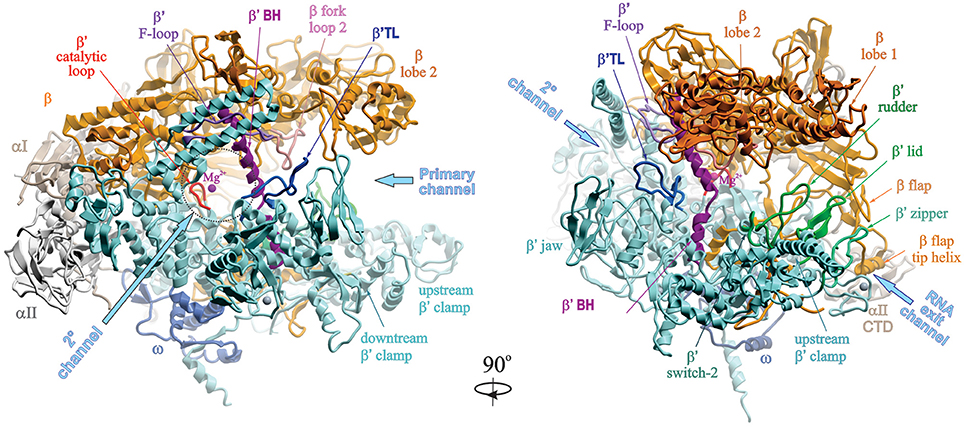
Bacterial RNA Polymerase-DNA Interaction—The Driving Force of Gene Expression and the Target for Drug Action

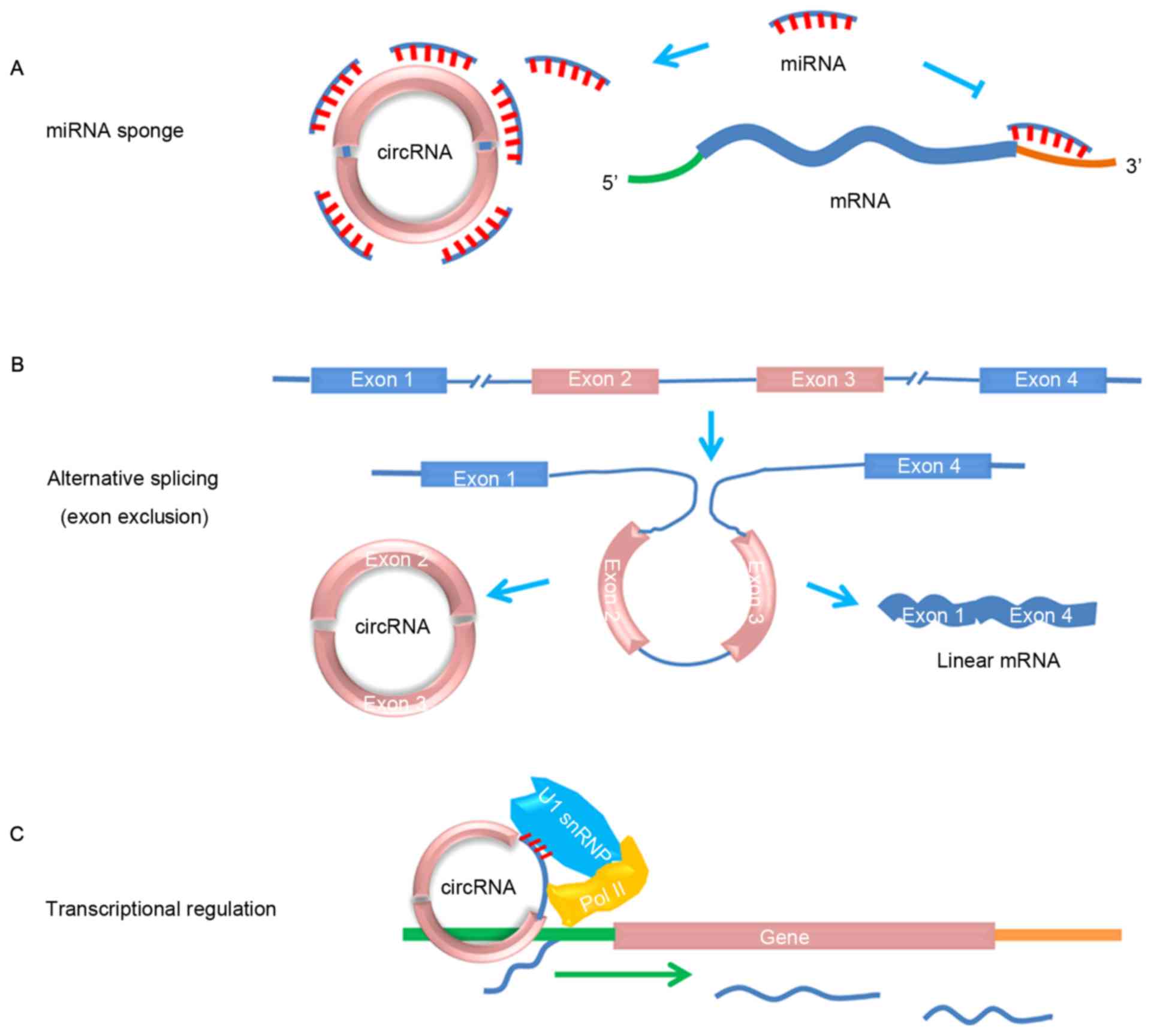
Oncology LettersPotential functions and implications of circular RNA in gastrointestinal cancer (Review)


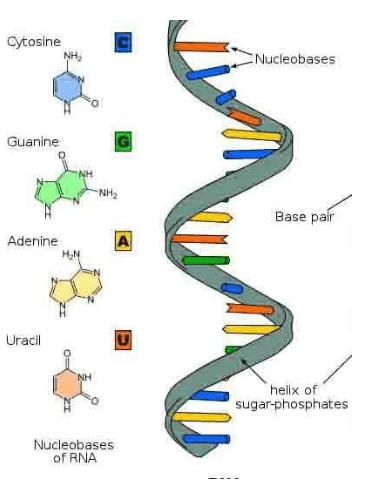
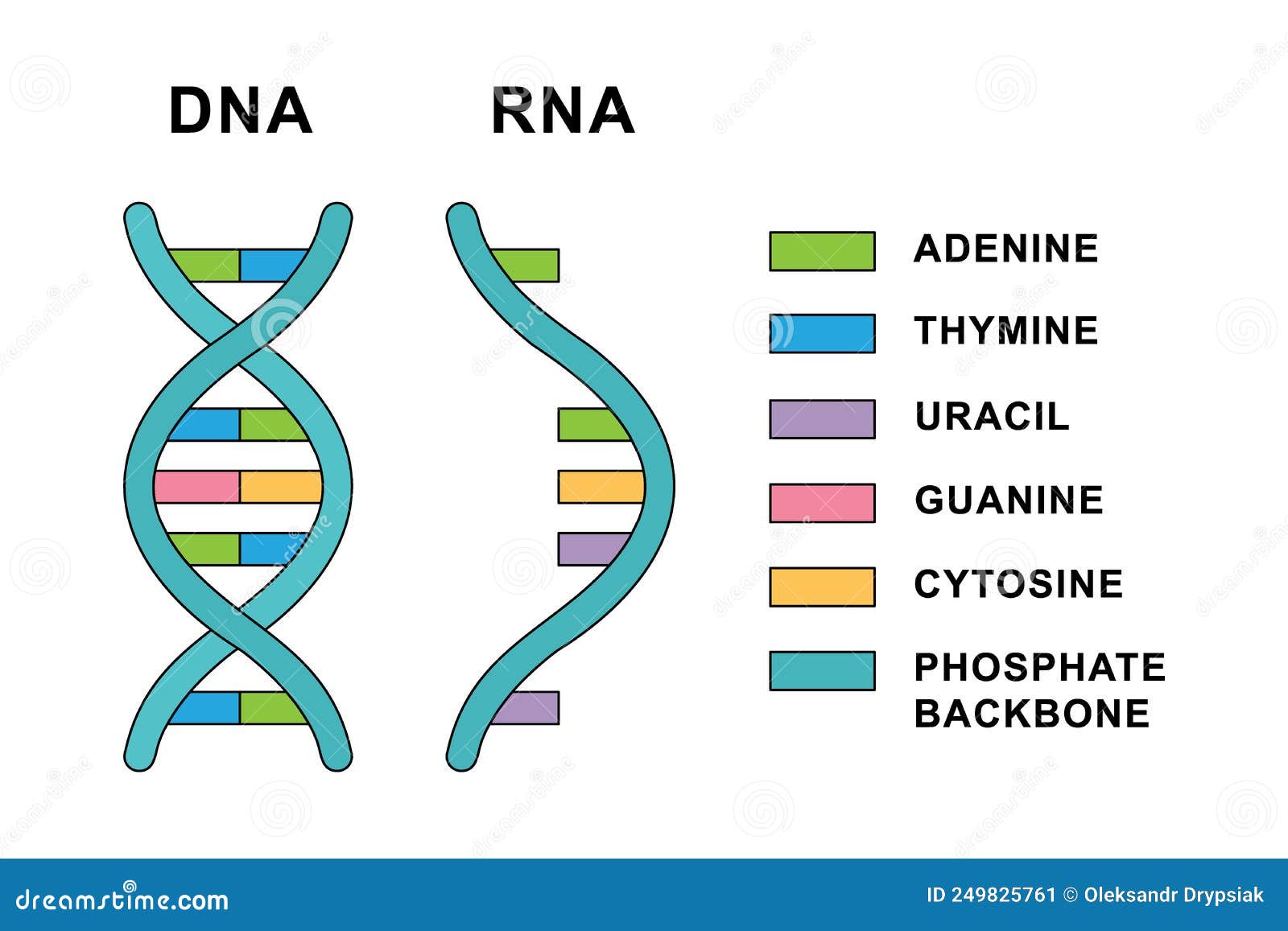
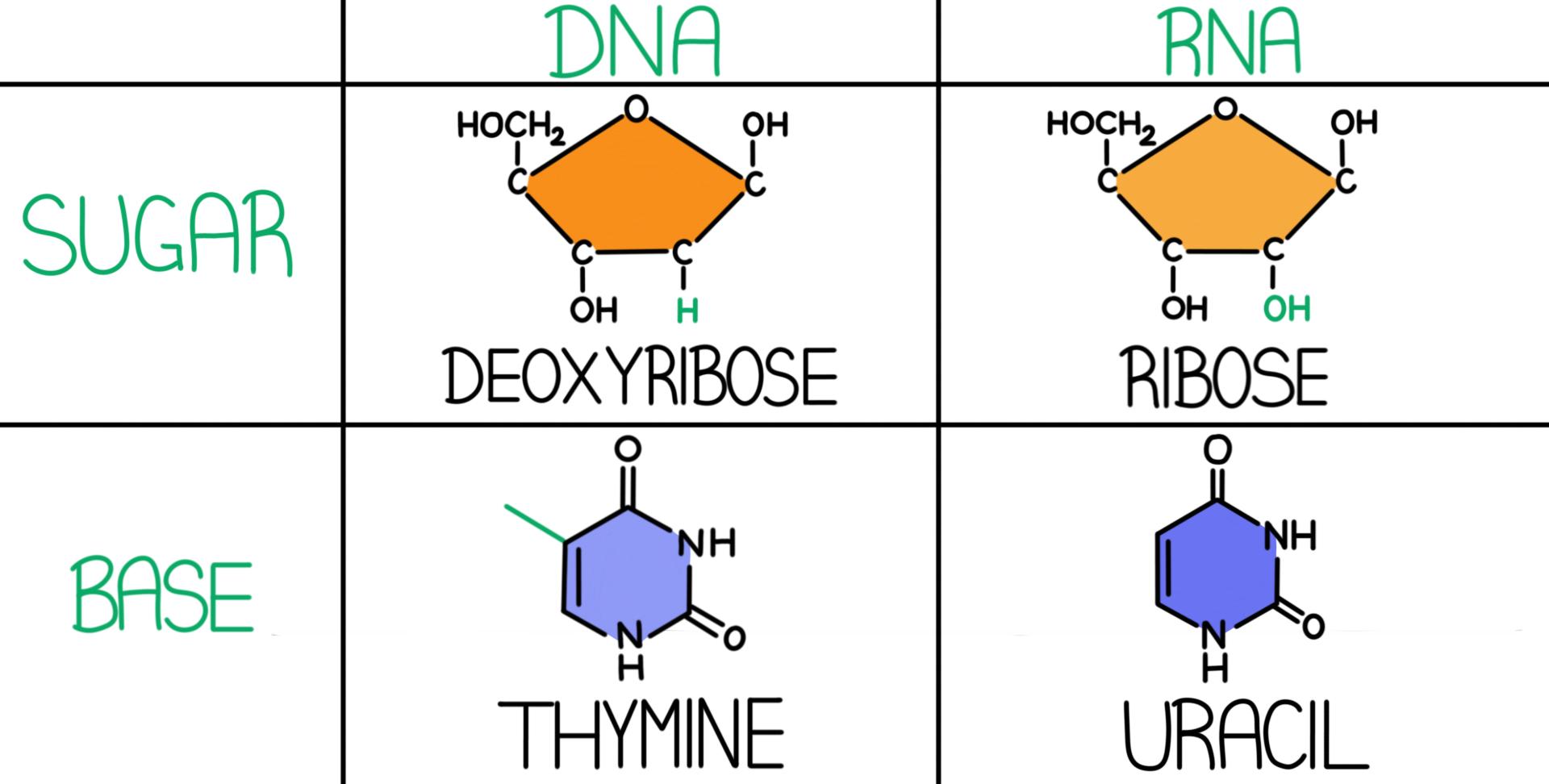
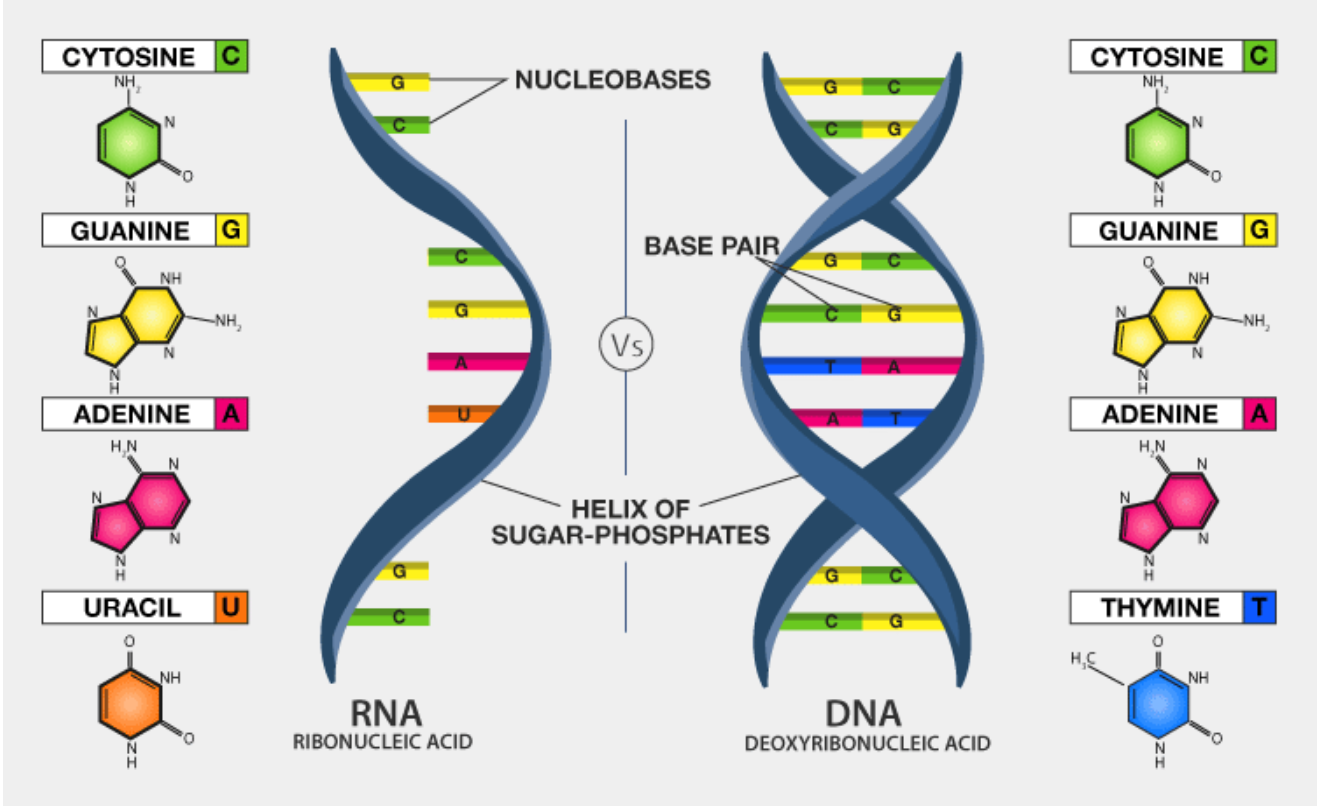
Structure of Ribonucleic acid and Deoxyribonucleic acid molecules. DNA and RNA nucleobases structure - cytosine, guanine. Structure of spirals Ribonucleic acid and Deoxyribonucleic acid molecules. DNA and RNA molecules with nucleobases structure - cytosine, guanine, adenine, uracil, thymine. Helical structure


Long non-coding RNA H19 promotes the proliferation and invasion of lung cancer cells and regulates the expression of E-cadherin, N-cadherin, and vimentin







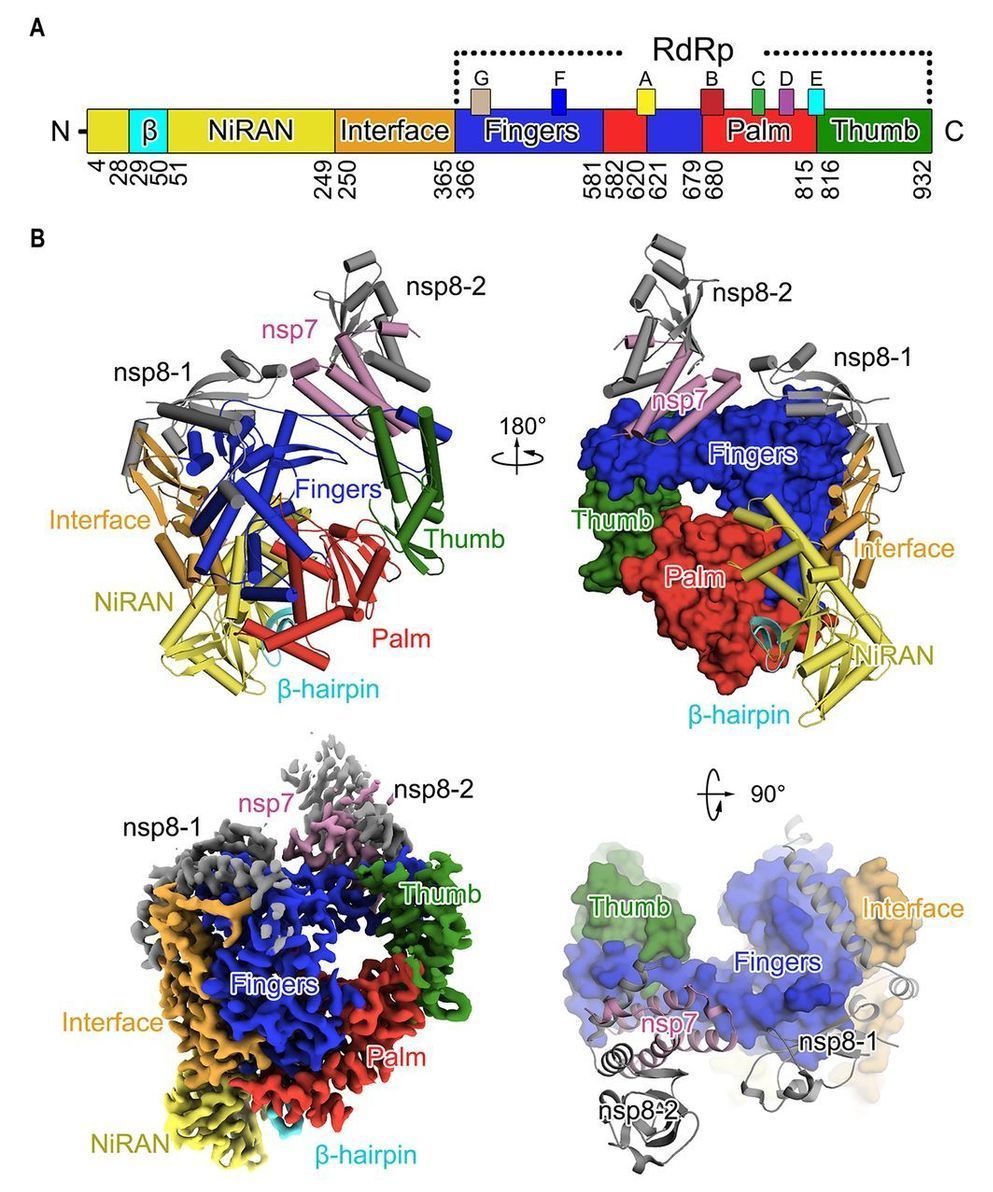


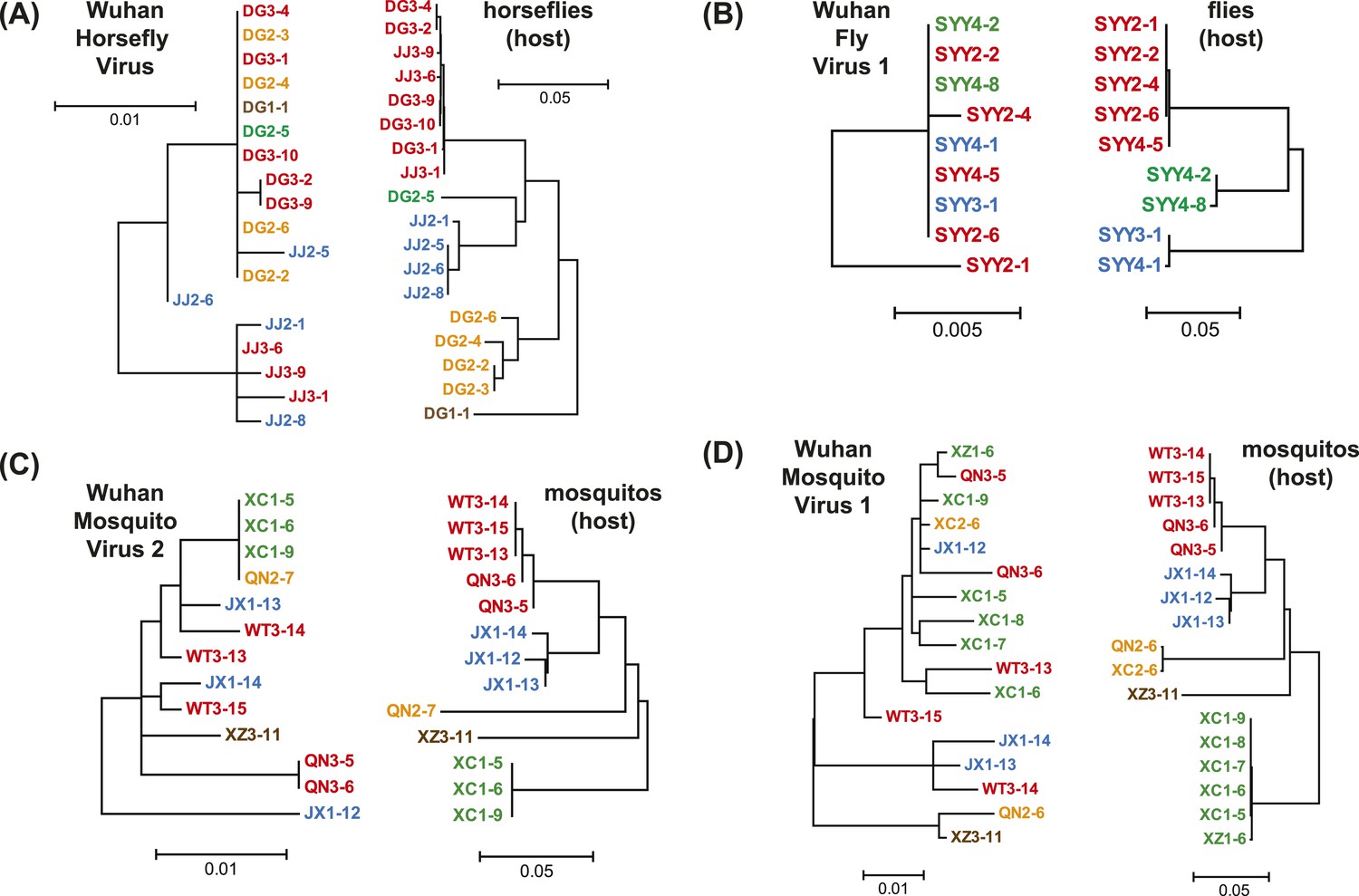
Unprecedented genomic diversity of RNA viruses in arthropods reveals the ancestry of negative-sense RNA viruses Phylogeny: Are arthropods at the heart of virus evolution? Characterisation of an Escherichia coli line that completely lacks ribonucleotide reduction yields insights into the evolution of parasitism and endosymbiosis Interactions between metabolism and growth can determine the co-existence of Staphylococcus aureus and Pseudomonas aeruginosa

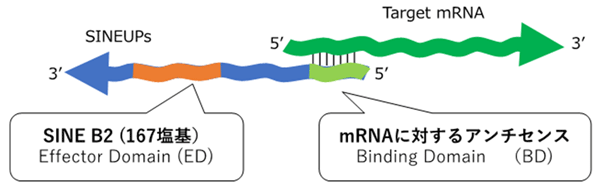



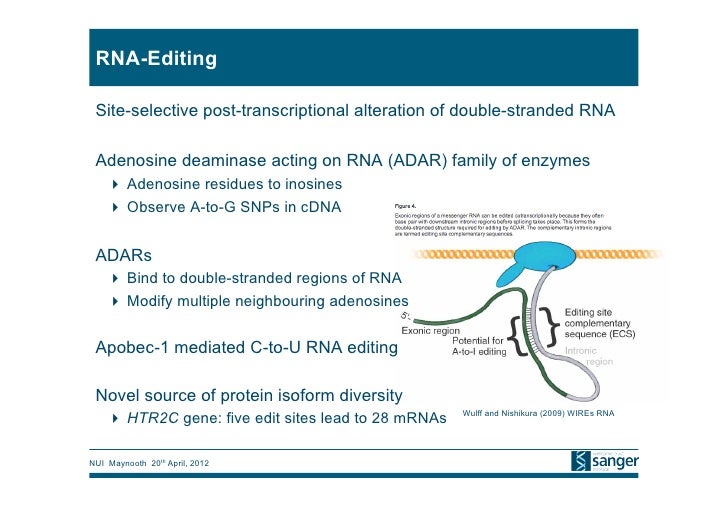
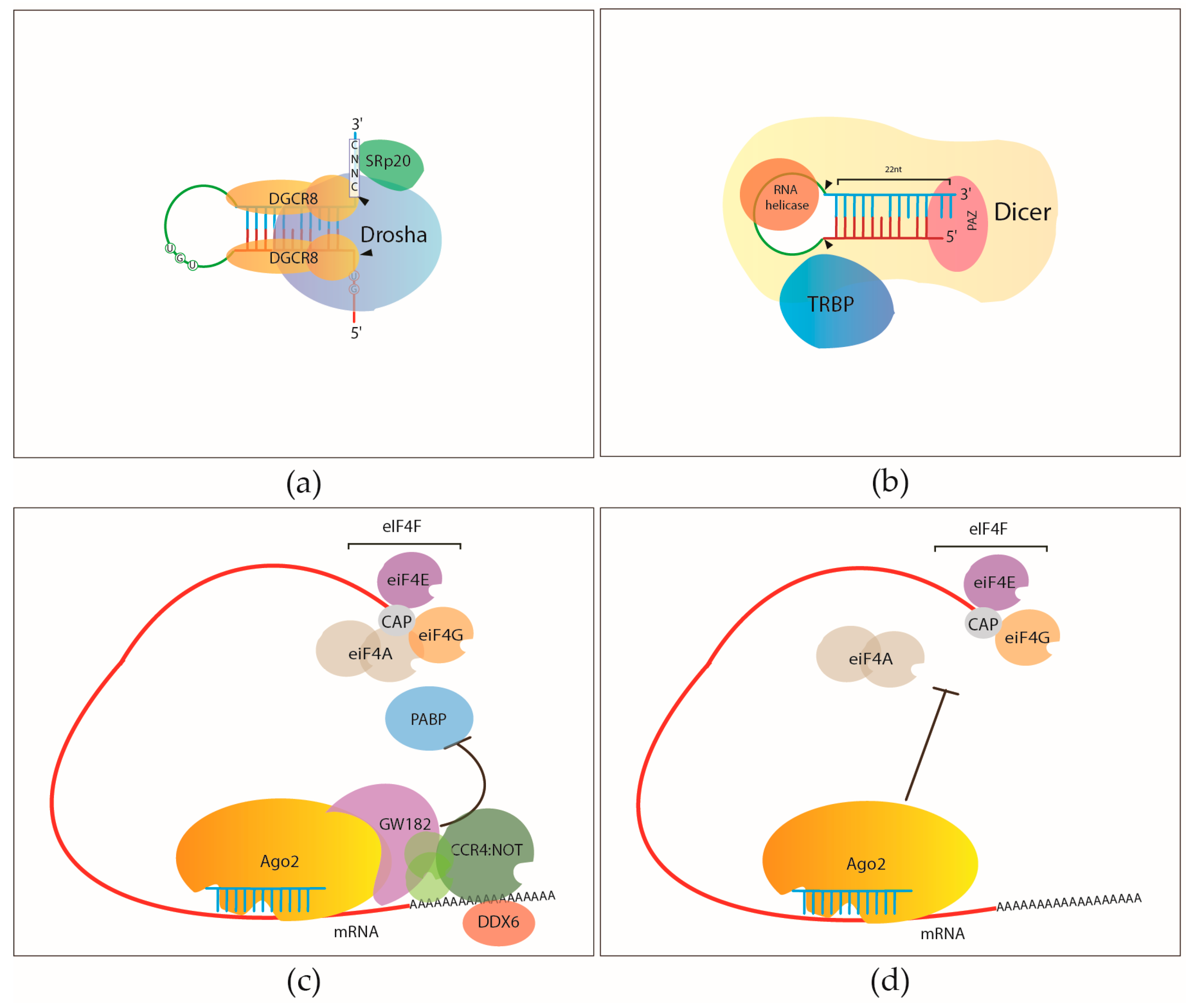
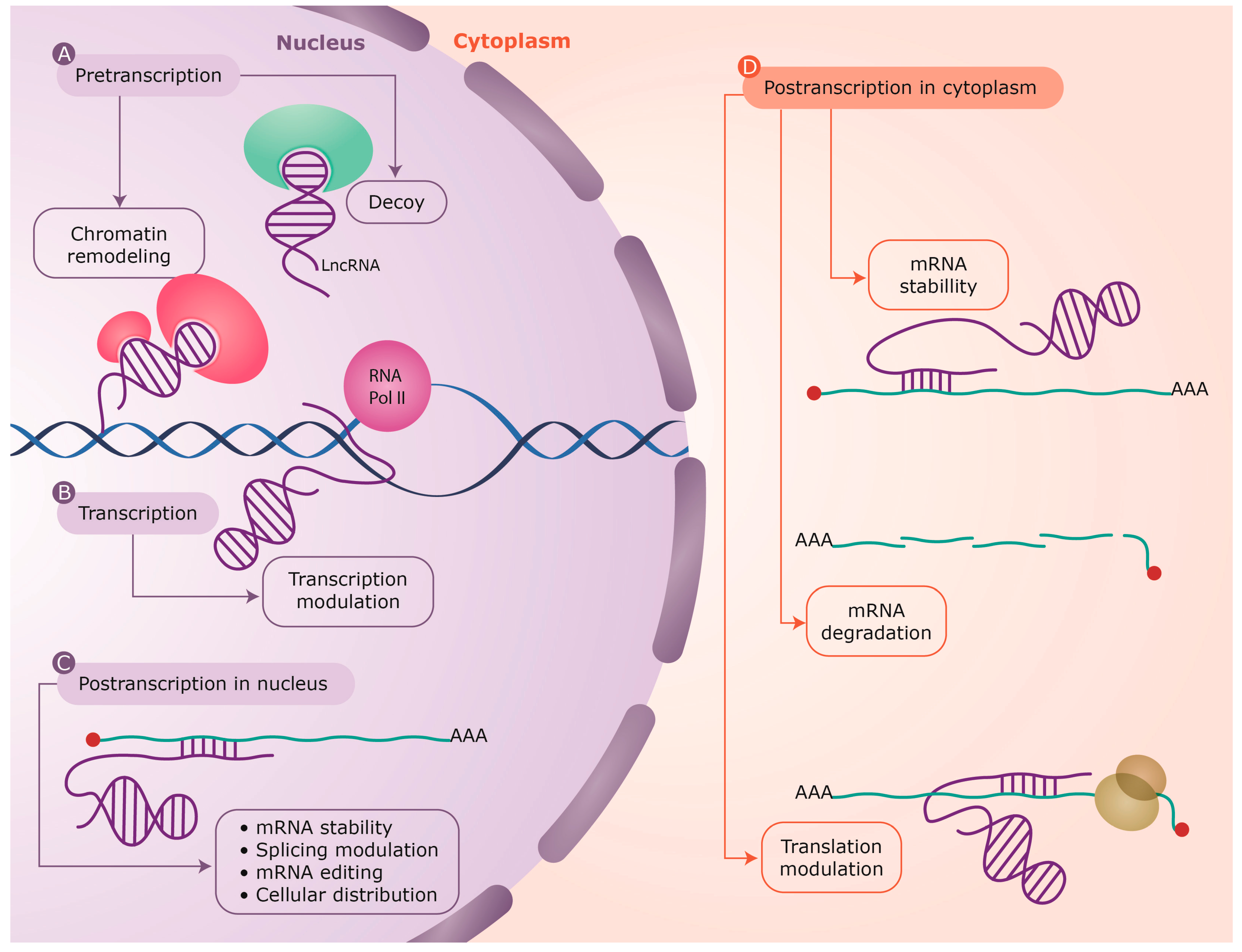
![RNA editing in the mammalian brain; One area that is known to require extensive proteindiversity is the brain. It is well known that pre-mRNA processing such asalternative splicing is widespread and highly regulated in the nervous system.Another way to alter the mRNA is by RNA editing. Adenosine to inosine (A-to-I)deamination is the most common type of RNA editing found in mammals and it iscatalyzed by adenosine deaminases that act on RNA (ADARs). There are twoenzymes found to be active in adenosine deamination, ADAR1 and ADAR2. These enzymesconvert A-to-I within double-stranded or highly structured RNA. Since inosineis recognized as guanosine by the cellular machinery, A-to-I editing has thepotential to change the code for translation. Site-selective A-to-I editing isa mechanism used to fine-tune the transcriptome and increase the variety ofexpressed protein isoforms, mainly found in the central nervous system (CNS).Besides amino acid changes, editing has been found to be able to influence theoutcome of splicing as well as other post-transcriptional events like 3’ endprocessing, stability, and transport (Figure 1). Thus, RNA editing by A-to-Imodification has the power to affect the proteome in many different ways. http://www.su.se/Apolipoprotein(apo)B circulates in two distinct forms, apoB100 and apoB48. Human liversecretes apoB100, the product of a large mRNA encoding 4564 residues. The smallintestine of all mammals secretes apoB48, which arises following C-to-Udeamination of a single cytidine base in the nuclear apoB transcript,introducing a translational stop codon. This process, referred to as apoB RNAediting, operates through a multicomponent enzyme complex that contains asingle catalytic subunit, apobec-1, in addition to other protein factors thathave yet to be cloned. ApoB RNA editing also exhibits stringent cis-actingrequirements that include both structural and sequence-specificelements—specifically efficiency elements that flank the minimal cassette, anAU-rich RNA context, and an 11-nucleotide mooring sequence—located in proximityto a suitably positioned (usually upstream) cytidine. C-to-U RNA editing maybecome unconstrained under circumstances where apobec-1 is overexpressed, inwhich case multiple cytidines in apoB RNA, as well as in other transcripts,undergo C-to-U editing. ApoB RNA editing is eliminated following targeting ofapobec-1, establishing that there is no genetic redundancy in this function.Under physiological circumstances, apoB RNA editing exhibits developmental, hormonal,and nutritional regulation, in some cases related to transcriptional regulationof apobec-1 mRNA. ApoB and the microsomal triglyceride transfer protein (MTP)are essential for the assembly and secretion of apoB-containing lipoproteins.MTP functions by transferring lipid to apoB during its translation and bytransporting triglycerides into the endoplasmic reticulum to form apoB-freelipid droplets. These droplets fuse with nascent apoB-containing particles toform mature, very low-density lipoproteins or chylomicrons. In cultured hepaticcells, lipid availability dictates the rate of apoB production. Unlipidated orunderlipidated forms of apoB are subjected to presecretory degradation, aprocess mediated by retrograde transport from the lumen of the endoplasmicreticulum to the cytosol, coupled with multiubquitination and proteasomaldegradation. Although control of lipid secretion in vivo is primarily achievedat the level of lipoprotein particle size, regulation of apoB production bypresecretory degradation may be relevant in some dyslipidemic states. Apolipoprotein B: mRNA Editing, Lipoprotein Assembly, and Pre secretoryDegradation; Annual Review of Nutrition.TermsRecognition of the edited nucleotide by the editingenzymes.a | APOBEC1 binds to APOB mRNA in the presence of ACF andcatalyses the C Udeamination of C6666 thatis positioned at the active site. According to one model, uracil is positionedat the pseudoactive site. It is not certain whether APOBEC1 binds as a dimer ormonomer when it is complexed with ACF. The cis-acting sequence elements are a MOORING SEQUENCE, and 5' and 3' efficiency elements.This is one of the many models of the configuration of the APOB-bindingsite. b | ADARs recognize duplex RNA that isformed between the editing site and the ECS that is often located in adownstream intron. The enzymes bind to the double-stranded (ds)RNA throughtheir dsRBDs and deaminate a specific adenosine to inosine. ACF, APOBEC1complementation factor; ADAR, adenosine deaminases that act on RNA; APOB,apolipoprotein B; APOBEC1, APOB mRNA-editingenzyme catalytic polypeptide 1; C, cytosine; ECS, editing site complementarysequence.; Liam P. Keegan, Angela Gallo & MaryA. O'Connell; www.Nature,comThe structural landscape of native Editosomesin African Trypanosomes Trypanosome RNA Editing: Simple Guide RNAFeatures Enhance U Deletion 100-Fold; Current understanding of U deletion andU insertion. RNA editing is catalyzed by a complex with seven majorpolypeptides, two of which correspond to distinct ligases (12, 29). U deletion and U insertion involvegRNA-directed endonuclease cleavage of the pre-mRNA (shown by an arrowhead),either 3′-U-exo or TUTase acting on the upstream fragment, and RNA ligaserejoining the mRNA (10, 17, 32). R, purine(s); Y, pyrimidine(s). Thesetwo forms of editing use distinct cleavage activities (11) and distinct ligase enzymes(Cruz-Reyes et al., unpublished), and the 3′-U-exo is not a reverseTUTase reaction (10, 29), which is indicated by the unequalsign. gRNAs have three main portions: a 5′ region to anchor the gRNA tothe mRNA just downstream of the editing site and direct the cleavage, a centralregion to guide the U deletions and insertions at mismatches in base pairingwith the pre-mRNA, and a 3′ oligo(U) that may tether the very purine-richupstream pre-mRNA. Many additional conserved gRNA features have also been noted(see the text). Dotted lines indicate potential base pairs that could serve ina ligation CatherineE. Huang, Jorge Cruz-Reyes, Alevtina G. Zhelonkina, Sean O'Hearn, ElizabethWirtz and Barbara Sollner-WebbInsertion/DeletionEditingWhen does ALS start? ADAR2-GluA2 hypothesisfor the etiology of sporadic ALS; Amyotrophic lateralsclerosis (ALS) is the most common adult-onset motor neuron disease. More than90% of ALS cases are sporadic, and the majority of sporadic ALS patients do notcarry mutations in genes causative of familial ALS; therefore, investigationspecifically targeting sporadic ALS is needed to discover the pathogenesis. Themotor neurons of sporadic ALS patients express unedited GluA2 mRNA at the Q/Rsite in a disease-specific and motor neuron-selective manner. GluA2 is asubunit of the AMPA receptor, and it has a regulatory role in theCa(2+)-permeability of the AMPA receptor after the genomic Q codon is replacedwith the R codon in mRNA by adenosine-inosine conversion, which is mediated byadenosine deaminase acting on RNA 2 (ADAR2). Therefore, ADAR2 activity may notbe sufficient to edit all GluA2 mRNA expressed in the motor neurons of ALSpatients. To investigate whether deficient ADAR2 activity plays pathogenicroles in sporadic ALS, we generated genetically modified mice (AR2) in whichthe ADAR2 gene was conditionally knocked out in the motor neurons. AR2 miceshowed an ALS-like phenotype with the death of ADAR2-lacking motor neurons.Notably, the motor neurons deficient in ADAR2 survived when they expressed onlyedited GluA2 in AR2/GluR-B(R/R) (AR2res) mice, in which the endogenous GluA2alleles were replaced by the GluR-B(R) allele that encoded edited GluA2. Inheterozygous AR2 mice with only one ADAR2 allele, approximately 20% of thespinal motor neurons expressed unedited GluA2 and underwent degeneration,indicating that half-normal ADAR2 activity is not sufficient to edit all GluA2expressed in motor neurons. It is likely therefore that the expression ofunedited GluA2 causes the death of motor neurons in sporadic ALS. Wehypothesize that a progressive downregulation of ADAR2 activity plays acritical role in the pathogenesis of sporadic ALS and that the pathologicalprocess commences when motor neurons express unedited GluA2. When does ALS start? ADAR2-GluA2 hypothesis for the etiology of sporadic ALS(PDF Download Available). Available from: https://www.researchgate.net/publication/51816954_When_does_ALS_start_ADAR2-GluA2_hypothesis_for_the_etiology_of_sporadic_ALS[accessed Jun 11, 2017].The Good and the Bad of Glutamate ReceptorRNA Editing; Glutamate receptors play a key role in excitatory synaptictransmission and plasticity in the central nervous system (CNS). Their channelproperties are largely dictated by the subunit composition of tetramericreceptors. Amino-3-hydroxy-5-methyl-4-isoxazolepropionic acid (AMPA) andkainate channels are assembled from GluA1–4 AMPA or GluK1–5 kainate receptorsubunits. However, their functional properties are highly modulated by apost-transcriptional mechanism called RNA editing. This process involves theenzymatic deamination of specific adenosines (A) into inosines (I) inpre-messenger RNA. This post-transcriptional modification leads to criticalamino acid substitutions in the receptor subunits, which induce profoundalterations of the channel properties. Three of the four AMPA and two of thefive kainate receptor subunits are subjected to RNA editing. This study reviewsthe advances in understanding the importance of glutamate receptor RNA editingin finely tuning glutamatergic neurotransmission under physiological conditionsand discusses the way in which the dis-regulation of RNA editing may beinvolved in neurological pathology. The Good and the Bad of Glutamate Receptor RNA Editing (PDF Download Available).Available from: https://www.researchgate.net/publication/309311484_The_Good_and_the_Bad_of_Glutamate_Receptor_RNA_Editing[accessed Jun 11, 2017]. Alice Filippini et al.](https://mol-biol4masters.masters.grkraj.org/html/RNA_Processing5-Editing_of_mRNA_Transcripts_files/image041.jpg)
RNA editing in the mammalian brain; One area that is known to require extensive proteindiversity is the brain. It is well known that pre-mRNA processing such asalternative splicing is widespread and highly regulated in the nervous system.Another way to alter the mRNA is by RNA editing. Adenosine to inosine (A-to-I)deamination is the most common type of RNA editing found in mammals and it iscatalyzed by adenosine deaminases that act on RNA (ADARs). There are twoenzymes found to be active in adenosine deamination, ADAR1 and ADAR2. These enzymesconvert A-to-I within double-stranded or highly structured RNA. Since inosineis recognized as guanosine by the cellular machinery, A-to-I editing has thepotential to change the code for translation. Site-selective A-to-I editing isa mechanism used to fine-tune the transcriptome and increase the variety ofexpressed protein isoforms, mainly found in the central nervous system (CNS).Besides amino acid changes, editing has been found to be able to influence theoutcome of splicing as well as other post-transcriptional events like 3’ endprocessing, stability, and transport (Figure 1). Thus, RNA editing by A-to-Imodification has the power to affect the proteome in many different ways. http://www.su.se/Apolipoprotein(apo)B circulates in two distinct forms, apoB100 and apoB48. Human liversecretes apoB100, the product of a large mRNA encoding 4564 residues. The smallintestine of all mammals secretes apoB48, which arises following C-to-Udeamination of a single cytidine base in the nuclear apoB transcript,introducing a translational stop codon. This process, referred to as apoB RNAediting, operates through a multicomponent enzyme complex that contains asingle catalytic subunit, apobec-1, in addition to other protein factors thathave yet to be cloned. ApoB RNA editing also exhibits stringent cis-actingrequirements that include both structural and sequence-specificelements—specifically efficiency elements that flank the minimal cassette, anAU-rich RNA context, and an 11-nucleotide mooring sequence—located in proximityto a suitably positioned (usually upstream) cytidine. C-to-U RNA editing maybecome unconstrained under circumstances where apobec-1 is overexpressed, inwhich case multiple cytidines in apoB RNA, as well as in other transcripts,undergo C-to-U editing. ApoB RNA editing is eliminated following targeting ofapobec-1, establishing that there is no genetic redundancy in this function.Under physiological circumstances, apoB RNA editing exhibits developmental, hormonal,and nutritional regulation, in some cases related to transcriptional regulationof apobec-1 mRNA. ApoB and the microsomal triglyceride transfer protein (MTP)are essential for the assembly and secretion of apoB-containing lipoproteins.MTP functions by transferring lipid to apoB during its translation and bytransporting triglycerides into the endoplasmic reticulum to form apoB-freelipid droplets. These droplets fuse with nascent apoB-containing particles toform mature, very low-density lipoproteins or chylomicrons. In cultured hepaticcells, lipid availability dictates the rate of apoB production. Unlipidated orunderlipidated forms of apoB are subjected to presecretory degradation, aprocess mediated by retrograde transport from the lumen of the endoplasmicreticulum to the cytosol, coupled with multiubquitination and proteasomaldegradation. Although control of lipid secretion in vivo is primarily achievedat the level of lipoprotein particle size, regulation of apoB production bypresecretory degradation may be relevant in some dyslipidemic states. Apolipoprotein B: mRNA Editing, Lipoprotein Assembly, and Pre secretoryDegradation; Annual Review of Nutrition.TermsRecognition of the edited nucleotide by the editingenzymes.a | APOBEC1 binds to APOB mRNA in the presence of ACF andcatalyses the C Udeamination of C6666 thatis positioned at the active site. According to one model, uracil is positionedat the pseudoactive site. It is not certain whether APOBEC1 binds as a dimer ormonomer when it is complexed with ACF. The cis-acting sequence elements are a MOORING SEQUENCE, and 5' and 3' efficiency elements.This is one of the many models of the configuration of the APOB-bindingsite. b | ADARs recognize duplex RNA that isformed between the editing site and the ECS that is often located in adownstream intron. The enzymes bind to the double-stranded (ds)RNA throughtheir dsRBDs and deaminate a specific adenosine to inosine. ACF, APOBEC1complementation factor; ADAR, adenosine deaminases that act on RNA; APOB,apolipoprotein B; APOBEC1, APOB mRNA-editingenzyme catalytic polypeptide 1; C, cytosine; ECS, editing site complementarysequence.; Liam P. Keegan, Angela Gallo & MaryA. O'Connell; www.Nature,comThe structural landscape of native Editosomesin African Trypanosomes Trypanosome RNA Editing: Simple Guide RNAFeatures Enhance U Deletion 100-Fold; Current understanding of U deletion andU insertion. RNA editing is catalyzed by a complex with seven majorpolypeptides, two of which correspond to distinct ligases (12, 29). U deletion and U insertion involvegRNA-directed endonuclease cleavage of the pre-mRNA (shown by an arrowhead),either 3′-U-exo or TUTase acting on the upstream fragment, and RNA ligaserejoining the mRNA (10, 17, 32). R, purine(s); Y, pyrimidine(s). Thesetwo forms of editing use distinct cleavage activities (11) and distinct ligase enzymes(Cruz-Reyes et al., unpublished), and the 3′-U-exo is not a reverseTUTase reaction (10, 29), which is indicated by the unequalsign. gRNAs have three main portions: a 5′ region to anchor the gRNA tothe mRNA just downstream of the editing site and direct the cleavage, a centralregion to guide the U deletions and insertions at mismatches in base pairingwith the pre-mRNA, and a 3′ oligo(U) that may tether the very purine-richupstream pre-mRNA. Many additional conserved gRNA features have also been noted(see the text). Dotted lines indicate potential base pairs that could serve ina ligation CatherineE. Huang, Jorge Cruz-Reyes, Alevtina G. Zhelonkina, Sean O'Hearn, ElizabethWirtz and Barbara Sollner-WebbInsertion/DeletionEditingWhen does ALS start? ADAR2-GluA2 hypothesisfor the etiology of sporadic ALS; Amyotrophic lateralsclerosis (ALS) is the most common adult-onset motor neuron disease. More than90% of ALS cases are sporadic, and the majority of sporadic ALS patients do notcarry mutations in genes causative of familial ALS; therefore, investigationspecifically targeting sporadic ALS is needed to discover the pathogenesis. Themotor neurons of sporadic ALS patients express unedited GluA2 mRNA at the Q/Rsite in a disease-specific and motor neuron-selective manner. GluA2 is asubunit of the AMPA receptor, and it has a regulatory role in theCa(2+)-permeability of the AMPA receptor after the genomic Q codon is replacedwith the R codon in mRNA by adenosine-inosine conversion, which is mediated byadenosine deaminase acting on RNA 2 (ADAR2). Therefore, ADAR2 activity may notbe sufficient to edit all GluA2 mRNA expressed in the motor neurons of ALSpatients. To investigate whether deficient ADAR2 activity plays pathogenicroles in sporadic ALS, we generated genetically modified mice (AR2) in whichthe ADAR2 gene was conditionally knocked out in the motor neurons. AR2 miceshowed an ALS-like phenotype with the death of ADAR2-lacking motor neurons.Notably, the motor neurons deficient in ADAR2 survived when they expressed onlyedited GluA2 in AR2/GluR-B(R/R) (AR2res) mice, in which the endogenous GluA2alleles were replaced by the GluR-B(R) allele that encoded edited GluA2. Inheterozygous AR2 mice with only one ADAR2 allele, approximately 20% of thespinal motor neurons expressed unedited GluA2 and underwent degeneration,indicating that half-normal ADAR2 activity is not sufficient to edit all GluA2expressed in motor neurons. It is likely therefore that the expression ofunedited GluA2 causes the death of motor neurons in sporadic ALS. Wehypothesize that a progressive downregulation of ADAR2 activity plays acritical role in the pathogenesis of sporadic ALS and that the pathologicalprocess commences when motor neurons express unedited GluA2. When does ALS start? ADAR2-GluA2 hypothesis for the etiology of sporadic ALS(PDF Download Available). Available from: https://www.researchgate.net/publication/51816954_When_does_ALS_start_ADAR2-GluA2_hypothesis_for_the_etiology_of_sporadic_ALS[accessed Jun 11, 2017].The Good and the Bad of Glutamate ReceptorRNA Editing; Glutamate receptors play a key role in excitatory synaptictransmission and plasticity in the central nervous system (CNS). Their channelproperties are largely dictated by the subunit composition of tetramericreceptors. Amino-3-hydroxy-5-methyl-4-isoxazolepropionic acid (AMPA) andkainate channels are assembled from GluA1–4 AMPA or GluK1–5 kainate receptorsubunits. However, their functional properties are highly modulated by apost-transcriptional mechanism called RNA editing. This process involves theenzymatic deamination of specific adenosines (A) into inosines (I) inpre-messenger RNA. This post-transcriptional modification leads to criticalamino acid substitutions in the receptor subunits, which induce profoundalterations of the channel properties. Three of the four AMPA and two of thefive kainate receptor subunits are subjected to RNA editing. This study reviewsthe advances in understanding the importance of glutamate receptor RNA editingin finely tuning glutamatergic neurotransmission under physiological conditionsand discusses the way in which the dis-regulation of RNA editing may beinvolved in neurological pathology. The Good and the Bad of Glutamate Receptor RNA Editing (PDF Download Available).Available from: https://www.researchgate.net/publication/309311484_The_Good_and_the_Bad_of_Glutamate_Receptor_RNA_Editing[accessed Jun 11, 2017]. Alice Filippini et al.